Em abril de 1953, James Watson e Francis Crick agitaram a comunidade científica com um elegante modelo de dupla-hélice para a estrutura do ácido desoxirribonucleico, ou DNA. A foto ao lado mostra o modelo de DNA que eles construíram utilizando folhas de metal e arame. Ao longo dos últimos 60 anos, esse modelo se tornou o ícone da biologia moderna.

Em abril de 1953, James Watson e Francis Crick agitaram a comunidade científica com um elegante modelo de dupla-hélice para a estrutura do ácido desoxirribonucleico, ou DNA. A foto ao lado mostra o modelo de DNA que eles construíram utilizando folhas de metal e arame. Ao longo dos últimos 60 anos, esse modelo se tornou o ícone da biologia moderna.
Entre os cientistas que se concentraram na descoberta da estrutura tridimensional do DNA estavam Linus Pauling, no Instituto de Tecnologia da Califórnia, Maurice Wilkins e Rosalind Franklin, no King’s College de Londres. Os primeiros a oferecer a resposta correta, no entanto, foram dois cientistas relativamente desconhecidos na época – o americano James Watson e o inglês Francis Crick.
Rosalind Franklin e a sua difração por raios-X do DNA. Franklin, uma cristalógrafa consumada, conduziu os experimentos cruciais que resultaram na fotografia que permitiu a Watson e Crick a dedução da estrutura em dupla-hélice do DNA.

Os fatores de hereditariedade de Gregor Mendel e os genes em cromossomos de Thomas Hunt Morgan são, na verdade, compostos por material genético. Em termos químicos, nosso conteúdo genético é o que herdamos de nossos progenitores. A hereditariedade é a molécula mais celebrada da atualidade.
De todas as moléculas da natureza, os ácidos nucleicos são inigualáveis na habilidade de controlar a própria replicação a partir de monômeros. De fato, a semelhança da prole com os pais baseia-se na replicação precisa e na sua transmissão de uma geração para a próxima. A informação hereditária controla o desenvolvimento de nossas características bioquímicas, anatômicas, fisiológicas e, até certo ponto, comportamentais.
A estrutura do DNA
No modelo do DNA, existem duas cadeias principais compostas por açúcar e fosfato, que são antiparalelas, ou seja, orientadas em direções opostas. Você pode imaginar essa estrutura como uma escada de cordas com degraus rígidos: as laterais da escada representam as cadeias de açúcar-fosfato, enquanto os degraus correspondem aos pares de bases nitrogenadas.
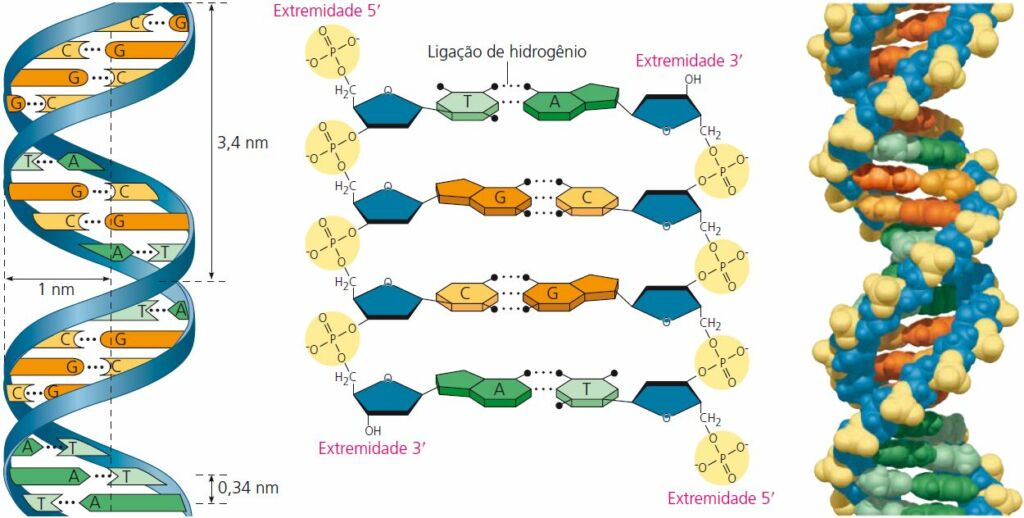
As bases nitrogenadas da dupla-hélice estão pareadas em combinações específicas: adenina (A) com timina (T), e guanina (G) com citosina (C). Adenina e guanina são purinas, bases nitrogenadas com dois anéis orgânicos: enquanto citosina e timina são bases nitrogenadas conhecidas como pirimidinas, que possuem apenas um anel.
Assim, as purinas (A e G) têm aproximadamente o dobro da largura das pirimidinas (C e T). Um par purina-purina é muito largo, e uma par pirimidina-pirimidina é muito estreito para os 2 nm de diâmetro da dupla-hélice. O pareamento purina-pirimidina, no entanto, resulta em um diâmetro uniforme.
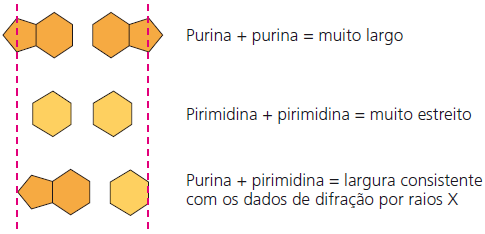
Assim, as purinas (A e G) têm aproximadamente o dobro da largura das pirimidinas (C e T). Um par purina-purina é muito largo, e uma par pirimidina-pirimidina é muito estreito para os 2 nm de diâmetro da dupla-hélice. O pareamento purina-pirimidina, no entanto, resulta em um diâmetro uniforme.
Cada um dos quatro monômeros de nucleotídeo do DNA é composto por uma base nitrogenada (T, A, C ou G), pelo açúcar desoxirribose (azul) e por um grupo fosfato (amarelo). O grupo fosfato de um nucleotídeo está ligado ao açúcar do próximo, formando a “cadeia principal” com alternância de grupos fosfato e açúcar, de onde se projetam as bases. A cadeia polinucleotídica tem orientação da extremidade 5′ (com grupo fosfato) para a extremidade 3′ (com grupo –OH do açúcar). 5′ e 3′ são aos números designados aos átomos de carbono do anel do açúcar.
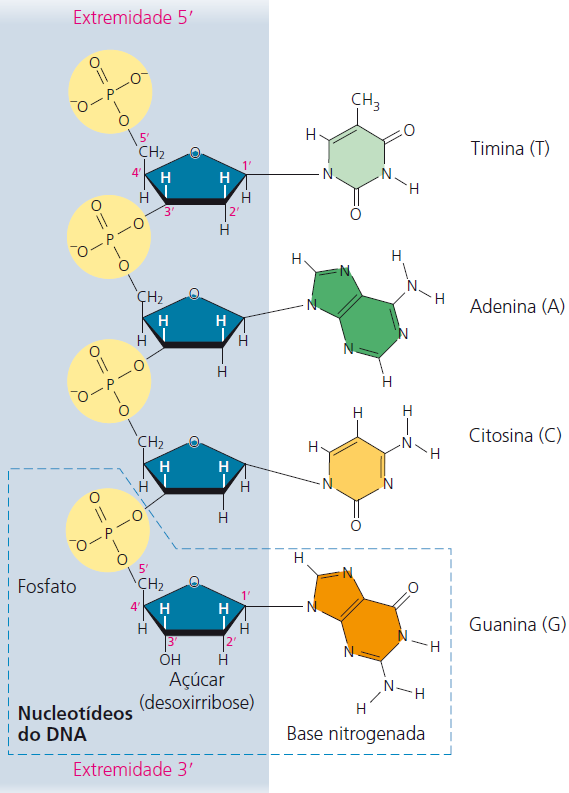
O bioquímico Erwin Chargaff chamou a atenção para a regularidade peculiar da proporção das bases dos nucleotídeos. Em cada espécie estudada, o número de adeninas e timinas era aproximadamente o mesmo; e o número de guaninas e citosinas era aproximadamente o mesmo. No ouriço-do-mar, por exemplo, as análises de Chargaff indicaram a presença da seguinte porcentagem de bases: A = 5 32,8%, T = 5 32,1%; G = 5 17,7% e C = 5 17,3%. As porcentagens não são exatamente as mesmas devido às limitações das técnicas utilizadas por Chargaff.
Como é a estrutura do DNA?
O modelo de replicação do DNA
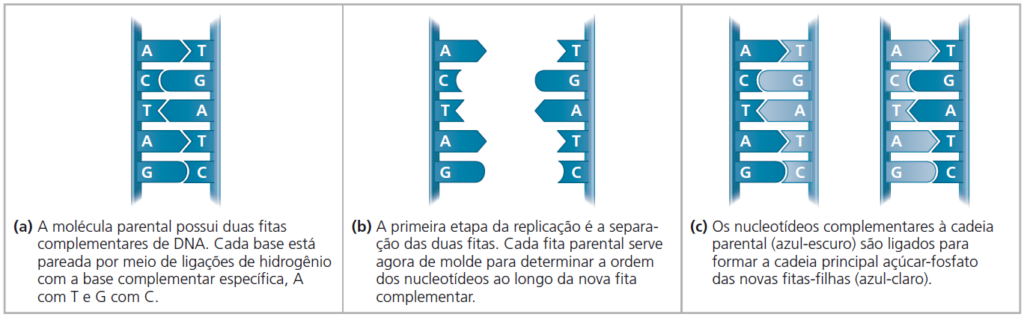
Quando a célula copia uma molécula de DNA, cada cadeia serve de molde para o ordenamento de nucleotídeos em uma nova fita complementar. Os nucleotídeos se alinham ao longo da fita-molde de acordo com as regras de pareamento e são ligados para formar novas fitas. Onde havia uma molécula de DNA dupla-fita no início do processo, haverá duas, cada uma sendo a cópia exata da molécula “parental”.
Vídeo
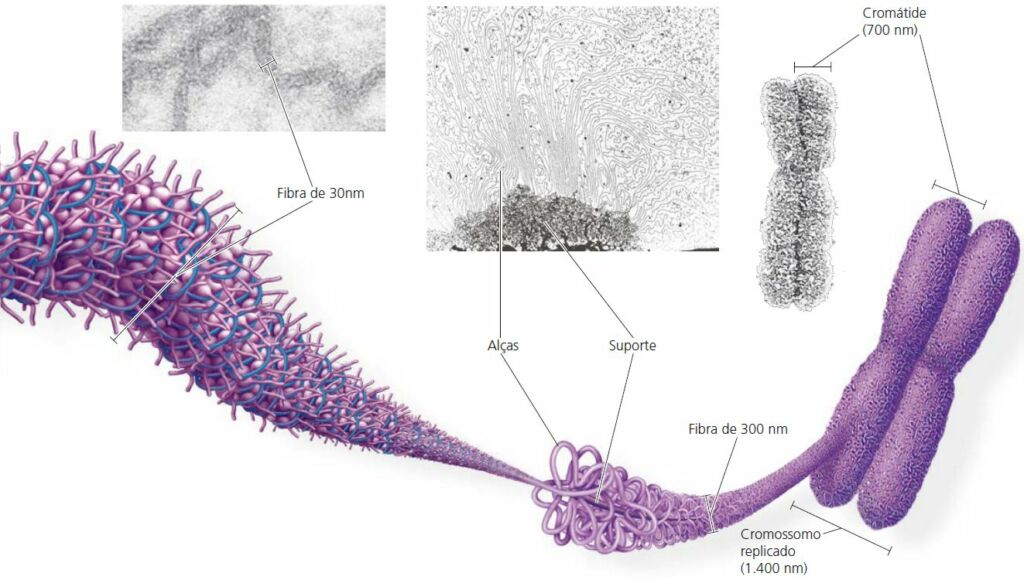
O cromossomo
Um cromossomo consiste em uma molécula de DNA empacotada com proteínas.
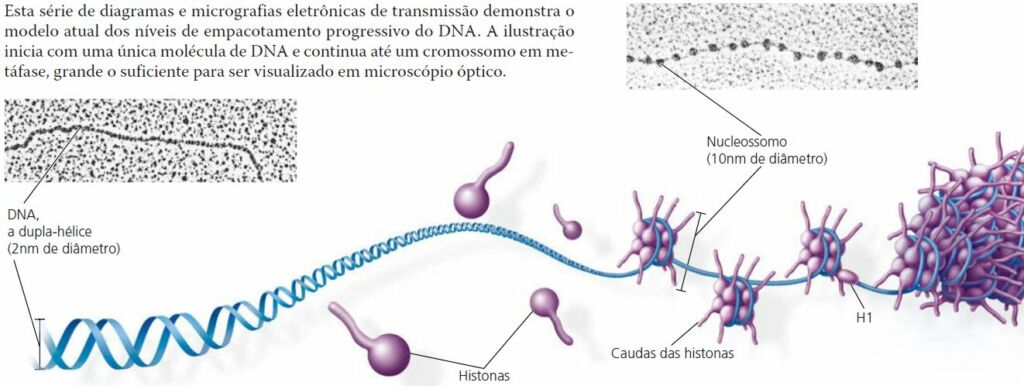
Cada cromossomo eucariótico contém uma única dupla-hélice linear com em média 1,5 x 108 pares de nucleotídeos. Essa é uma enorme quantidade em relação à extensão do cromossomo condensado. Se fosse completamente espichada, alcançaria 4 cm de comprimento, milhares de vezes o diâmetro do núcleo da célula – e isso sem considerar os demais 45 cromossomos humanos!
Proteínas chamadas histonas são responsáveis pelo primeiro nível de empacotamento. Apesar de cada histona ser pequena – com cerca de 100 aminoácidos -, a massa total de histonas na cromatina é quase igual à massa de material genético. Mais de 20% dos aminoácidos das histonas apresentam carga positiva (lisina ou arginina) e se ligam fortemente ao material.
Em micrografias eletrônicas, a cromatina desenrolada apresenta diâmetro de 10 nm. Esta cromatina lembra a aparência de um colar de contas. Cada “conta” é um nucleossomo, a unidade básica do empacotamento do DNA.
No ciclo celular, as histonas se dissociam do DNA apenas brevemente durante a sua replicação. Geralmente, elas também o fazem durante a transcrição, outro processo que requer o acesso ao DNA pelo processo molecular da célula.
Cromatina enovelada por histonas
Referência
As figuras apresentadas nesta matéria foram obtidas do recurso eletrônico Biologia de Campbell (REECE et al., 2015).
REECE, Jane B. et al. Biologia de Campbell [recurso eletrônico]. Tradução: Anne D. Villela et al.; revisão técnica: Denise Cantarelli Machado, Gaby Renard, Paulo Luiz de Oliveira. 10. ed. Porto Alegre: Artmed, 2015.